東アジアのナラ類とナラ林
鳥取大学乾燥地研究センター 山中典和
ナラ類をコナラ属に属する落葉性の種とすると、わが国に生育するナラ類はクヌギ節のクヌギとアベマキ、コナラ節のカシワ、ミズナラ、コナラ、ナラガシワの6種を挙げることが出来る。これらのナラ類はいずれも日本の温帯から暖帯の重要な構成要素であり、林業上有用な樹種である。このためわが国では古くから多くの研究が行われてきている。ちなみに森林総研のfolisで検索するとコナラというキーワードで814個、ミズナラで704個の文献がヒットする(10月4日現在)。
しかし、これほど多くの研究が行われており、日本人になじんでいるナラ類だが、その分布をみると、日本に特有の種は1種もない。日本に見られる6種すべてが中国大陸にその分布の中心を持っており、東アジアの暖帯域から温帯域を通じて重要な森林の構成種となっているのである。日本から飛び出し、東アジアの視点からナラ類を考えると、日本で見慣れている生態とはまた違った側面が見えてくる。
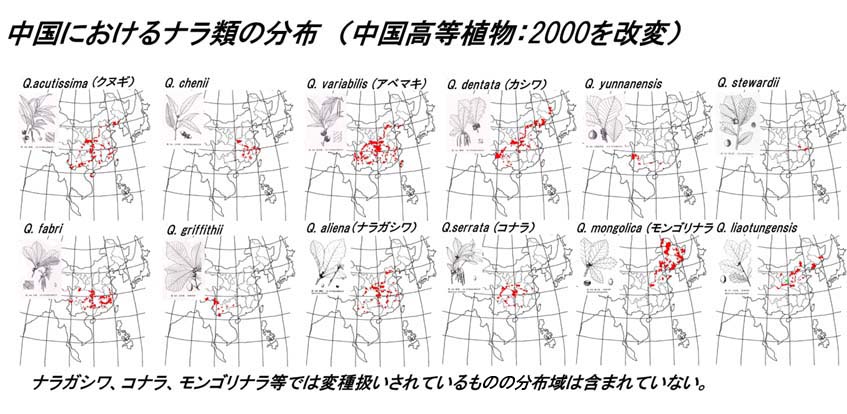
下に、2000年発行の中国高等植物をもとにした、中国でのナラ12種の分布域を示す。
中国のナラ類は研究者により若干の見解の相違はあるが、ほぼこの12種と考えてよいと思われる。朝鮮半島や沿海州等にもこれ以外の種はないようである。
中国のナラ類のうちクヌギ節はQ.accutissima (クヌギ)、Q.variabilis (アベマキ)、Q.cheniiの3種である。日本におけるクヌギ、アベマキの分布はどこまでが人為で、どこまでが自然分布か不明な点が多いが、中国での分布は非常に広く、黄河中下流域から長江流域にかけての暖帯域を中心として分布する。クヌギでは北は遼寧省が限界で、南は海南島まで幅広い緯度範囲で生育している。アベマキもクヌギとほぼ同様の分布域を示す。中国で、アベマキとクヌギが実際どのようなすみわけを行っているのか不明な点が多く、興味深い。中国樹木誌によるとアベマキとクヌギは混交する場合もあるようである。出現様式はマツ(馬尾松)等と混交することが多いようであるが、小面積の純林を作る場合もあるとされる。Q.cheniiはクヌギに似た種でクヌギよりも殻斗が小さい。分布はクヌギやアベマキに比べ狭く、北は河南省から南は福建省までみられる。
コナラ節はQ. dentata (カシワ)、Q.yunnanensis、Q. stewardii、Q. fabri、Q.Griffithii、Q. aliena (ナラガシワ)、Q. serrata (コナラ)、 Q. mongolica(モンゴリナラ)、Q. liaotungensis の9種である。
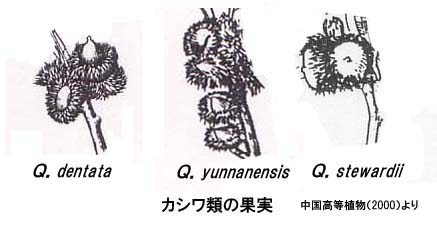
このうちQ. dentata (カシワ)、Q.yunnanensis、Q. stewardii は比較的近い種で、殻斗の形態が似ている。総苞片が長く伸び、直立または反曲することで、これら以外の6種と区別される。
カシワは中国国内の最北である黒竜江省まで分布し、ロシアの沿海州にも生育する等、寒さに強い種である。その反面、南は台湾まで分布が見られ、幅広い温度域に生育する。日当たり良い斜面の広葉樹林中や、マツ-ナラ林中に成育するようである。雑種としてQ. x hopeiensis や Q.x fangshanensis が記録されているが、この2雑種は中国樹木誌では独立種として取り扱われている。
Q.yunnanensisは分布が、四川や貴州、広東、広西、湖北の一部に限られており、海抜1000m_2600mの山地マツ林や広葉樹林中に出現する。中国樹木誌ではカシワの変種にされている。
Q.stewardii は安徽、浙江、江西、湖北省の海抜1000_1750mに分布する種で、分布域は狭い。安徽省の大別山では常緑カシであるQuercus glauca (アラカシ)等と混交林をつくっている様子である。
次に殻斗の総苞片が短く、全体が殻斗にくっついているタイプがQ. fabri、Q.Griffithii、Q. aliena (ナラガシワ)、Q. serrata (コナラ)、 Q. mongolica(モンゴリナラ)、Q. liaotungensisの6種である。
Q. fabriは、淮河以南、長江流域及び華南、西南を中心に分布する種で、主に海抜1900m以下の山地に生育する。Q.griffithiiは長さ20cm、大きなもので30cmに達する大きな葉を持つ種である。中国では雲南や貴州、四川等の西南部で海抜700_2800mの山地に生育し、インドやスリランカまで分布する。この2種は日本に分布しない。
ナラガシワ(Q. aliena )は日本では本州(山形、群馬、北陸、東京都以西)、四国、九州、に分布する種である。東アジアでの分布も広く、朝鮮、溝州、中国、印度支郡、タイなどの温帯、暖帯に広く分布する。中国でも暖帯域を中心とした分布をみせる。中国高等植物(2000)では変種として var. acuteserrata を認めている。
コナラ(Q. serrata)も分布が広い種であり、中国では長江中流域を中心とした分布をしめす。しかし、中国では変種として var. brevipetiolata (タイワンコナラ)を区別しており、この変種を含めるとその分布は上の地図に示された分布に加え、遼寧南部、山東省から南東の浙江、福建、台湾まで広がり、非常に広大な範囲になる。しかし、分布の中心は主に暖帯域となるようであり、あまり寒い地域には分布しない。

モンゴリナラの生態は日本で見慣れているミズナラと共通することも多く、中国東北部では伐採跡地や火事後などにモンゴリナラの一斉林を良く見かける。しかし、日本におけるミズナラと決定的に異なるのは、日本の冷温帯には圧倒的な優占種となりうる極相種のブナが存在するのに対し、中国には日本のブナに相当する種がいないことである。日本のミズナラ林はやがてブナ林へと移り変わり、ブナ林の中に単木的にミズナラの大木が混交する状態になることが多い。私が調査を行っている中国東北部の遼寧省東部ではブナの代わりに、サクラ類や、カエデ類、ボダイジュ類等々多様な樹種と混交したモンゴリナラニ次林をみることができる。このような林はやがてチョウセンゴヨウマツの混交した多様性の高い針広混交林へ移行すると考えられている。
もう一つ中国のモンゴリナラと日本のミズナラで異なる点は、その生育環境としての降水量である。日本の場合は全国どこにいっても樹木の生育に有り余る降水量が確保されているが、中国では乾燥地帯から湿潤地帯まで幅広い降水量傾度が存在する。モンゴリナラは年降水量1000mmを超える地域にも生育する反面、年降水量が500mmを切る森林の生育限界付近でも生育している。このような半乾燥地域では砂漠化が進行し、本来の生態系を窺い知る林分はほとんどないが、わずかに残された林分では、モンゴリナラを主体とし、少数ボダイジュ類や、カエデ類の混交が見られる。また油松(マンシュウクロマツ)とモンゴリナラの混交林もみられるが、斜面方位によるマツとナラの棲み分けもありそうで、今後の研究課題でもある。いずれにせよ中国東北部の半乾燥地帯ではモンゴリナラは極相を構成する重要な構成要素のようであり、同じ中国東北部の湿潤地帯に出現するモンゴリナラとは生態的地位や更新様式に違いが見られる。
モンゴリナラとよく似た分布傾向を見せるのがリョウトウナラ(Q. liaotungensis)である。
リョウトウナラはモンゴリナラほど北まで分布することはないが黄河流域の北側を中心とした中国の冷温帯域に分布し、一斉林を形成したり、油松との混交林分を形成したりする。さらにモンゴリナラ同様リョウトウナラは乾燥に強く、私が調査している中国の黄土高原では森林限界ぎりぎりの降水量450mm前後の地域までリョウトウナラの林分を見ることができる。しかしこの付近でも砂漠化による植生破壊が進み、森林はほとんど見られず、植林樹種としてニセアカシア等が多く植えられている。砂漠化防止のための乾燥地域での植林ではマツ類や外来樹種の使用が多いが、今後は在来植生を用いて生態系の修復も考えなくてはならず、リョウトウナラは黄土高原における生態系修復の鍵となる樹種と考えられる。

以上述べてきたように、ナラ類は日本のみならず東アジア全域に分布し、東アジア各地の生態系を考える際に非常に重要な位置を占めている。しかし、現実問題として、東アジアという視点からみたナラ類の生態、生理といった基礎的情報は驚くほど少ない。日本のナラ類も東アジアという広い分布域の中での位置付けをしてゆくことが必要であり、そこからまたナラ類の興味深い生態、生理特性が明らかになるものと思われる。
付録:中国ナラ類の検索表____中国高等植物(2000)より
1.落葉高木
2.葉は刺芒状の鋸歯、殻斗の総苞片は線形、あるいは短銛形の総苞片を兼ね備える
3葉の両面無毛あるいは下面脈上に毛がある。樹皮のコルク層は発達しない。若枝は柔毛あり。
4.殻斗は径2-4cm、果実は卵円形又は楕円形で径1.5-2cm____1.Q.accutissima (クヌギ)
4.殻斗は径約1.5cm、果実は楕円形、径1.3-1.5cm___________2.Q.chenii
3.葉の下面は密に灰白色の星状毛に被われる。樹皮はコルク層が発達、若枝無毛。
_________________________3.Q.variabilis (アベマキ)
2.葉は粗鋸歯又は波状鋸歯、殻斗の総苞片は被針形、三角形又は鱗片状。
5.殻斗の総苞片は被針形、長さ4mm以上で直立又は反曲する
6.葉の下面は密に星状毛で被われる。
7.殻斗の総苞片は長さ約1cmで紅褐色、褐色の絹毛がある。内面無毛、直立又は反曲。
___________________________4.Q. dentata (カシワ)
7.殻斗の総苞片長約8mm、灰黄又は褐色、灰色の絹毛がある。直立又は開展。
___________________________5. Q.yunnanensis
6.葉の下面無毛又は毛がある
8.葉柄長1-2cm、側脈9-12対 _______________4‘.Q. x fangshanensis
8.葉はほとんど無柄
9.側脈5-8(-10)対、実は楕円形_______________4“. Q. x hopeiensis
9.側脈10-16対、実は球形に近い_______________6.Q. stewardii
5.殻斗の総苞片は三角形、鱗片状又は卵状被針形、長さ4mm以下で、全体が殻斗にくっつく。
10.葉の下面星状毛を有す、また単毛を混じる。
11.小枝及び葉柄は密に毛で被われる。殻斗総苞片は褐色。
12.葉長15cmに達する。側脈8-12対、葉柄長3-5mm______7.Q. fabri
12.葉長20(-30)cmに達する。側脈12-18対、葉柄長0.5-1mm___8.Q.Griffithii
11.小枝ほぼ無毛、葉柄無毛、殻斗の総苞片は灰白色で短柔毛がある
13.葉は波状鋸歯_______________9. Q. aliena (ナラガシワ)
13.葉の鋸歯は粗く大きい、鋸歯の先端は鋭く尖り、内曲する。________________________________9’ Q. aliena var. acuteserrata
10.葉下面無毛、又はほぼ無毛
14.葉は腺歯
15.葉柄長1_3cm、葉は枝上に散生_________10. Q. serrata (コナラ)
15.葉柄長2_5mm、葉は常に枝先に輪生__10’.Q. serrata var.brevipetiolata
14.葉は鋸歯縁、鋸歯の先端には腺点はない。
16.葉柄長1_1.3cm________________9.Q. aliena (ナラガシワ)
16.葉柄長2_8mm
17.殻斗下部の総苞片には瘤状の突起がある
18.葉は鈍鋸歯又は粗鋸歯、側脈7_11対_-11.Q. mongolica(モンゴリナラ)
18.葉は内曲する粗鋸歯、側脈14_18対、比較的細長い。
______11’. Q. mongolica var.grosseserrata (ミズナラ)
17.殻斗総苞片は扁平、稀に下部やや厚い___12.Q. wutaishanica (liaotungensis)
(無断転載はお断りします)